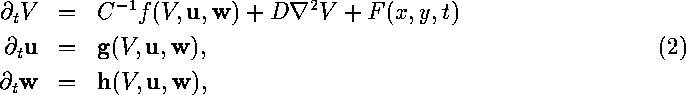The Oxsoft equations [9] summarise the results of
extensive voltage clamp and ion flux investigations on mammalian
ventricular tissue and cells, and provide a model for the membrane
currents resulting from voltage-dependent gated, and leakage,
conductances; active transmembrane exchanges; and intracellular ion
concentration changes, and ![]() release and sequestration from the
sarcoplasmic reticulum in a single ventricular
myocyte. In this paper, we use the model of guinea pig ventricular cell
from the Oxsoft family, the basic ideas of which can be found in
[10], the OGPV model. The same model is used in
[19], where it is described in more detail.
release and sequestration from the
sarcoplasmic reticulum in a single ventricular
myocyte. In this paper, we use the model of guinea pig ventricular cell
from the Oxsoft family, the basic ideas of which can be found in
[10], the OGPV model. The same model is used in
[19], where it is described in more detail.
For a single isopotential ventricular cell the model is in the form of a system of ordinary differential equations, which can be shortly written in the form
where V=V(t) is the transmembrane voltage, C is cell membrane
capacitance, f is transmembrane current per cell, vector ![]() describes the fast gating variables, and vector
describes the fast gating variables, and vector ![]() comprises slow gating variables and intra- and
extra-cellular ionic concentrations, and
comprises slow gating variables and intra- and
extra-cellular ionic concentrations, and ![]() and
and ![]() describe their kinetics. Action potential solutions of this model and
their rate dependence are illustrated in Fig. 1(a-c).
describe their kinetics. Action potential solutions of this model and
their rate dependence are illustrated in Fig. 1(a-c).
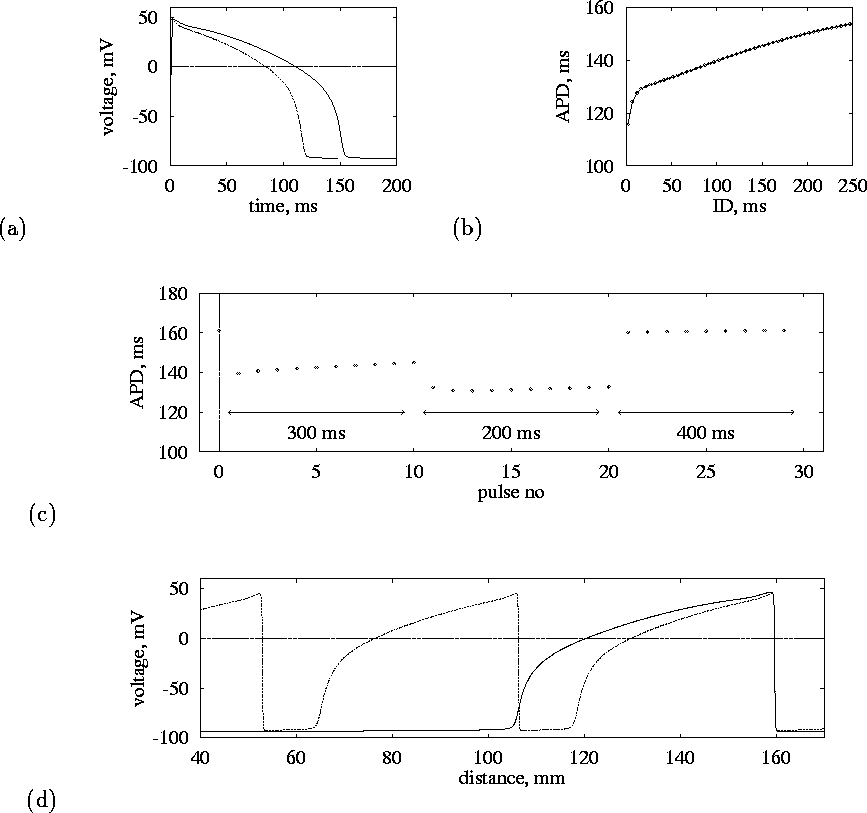
Figure 1:
The OGPV model.
(a) Action potential for normal ( ![]() , solid line) and high
(
, solid line) and high
( ![]() , dashed line) pacing rates.
(b) Restitution curve, action potential duration vs diastolic interval,
measured at 95% repolarisation (
, dashed line) pacing rates.
(b) Restitution curve, action potential duration vs diastolic interval,
measured at 95% repolarisation ( ![]() ).
(c) Variations of APD during pacing with changing period (shown by
arrows).
(d) Voltage profiles of propagating pulses, at normal (
).
(c) Variations of APD during pacing with changing period (shown by
arrows).
(d) Voltage profiles of propagating pulses, at normal ( ![]() ,
solid line) and high (
,
solid line) and high ( ![]() , dashed line) pacing
rates.
, dashed line) pacing
rates.
This model was incorporated into a partial differential equation model for an excitable medium in the plane (x,y)
where D is the diffusion coefficient for V, ![]() is the
Laplacian operator
is the
Laplacian operator ![]() and F(x,y,t) is a
time and space dependent forcing that models external electric current
applied to the tissue; as written in (2) it has the dimensionality
of voltage over time, and can be rescaled to current units via the
value of
and F(x,y,t) is a
time and space dependent forcing that models external electric current
applied to the tissue; as written in (2) it has the dimensionality
of voltage over time, and can be rescaled to current units via the
value of ![]() . This value of whole cell membrane
capacitance is the value used in the OGPV model; assuming the specific
membrane capacitance of
. This value of whole cell membrane
capacitance is the value used in the OGPV model; assuming the specific
membrane capacitance of ![]() this corresponds to
the membrane area of
this corresponds to
the membrane area of ![]() , which is about twice more than could be
deduced from cell sizes assumed in the OGPV model, radius of
, which is about twice more than could be
deduced from cell sizes assumed in the OGPV model, radius of
![]() and length of
and length of ![]() if the cell is to be considered as a perfect
cylinder.
The diffusion coefficient
if the cell is to be considered as a perfect
cylinder.
The diffusion coefficient ![]() was chosen
to give a conduction velocity for a solitary plane wave along one of
the coordinate axes of
was chosen
to give a conduction velocity for a solitary plane wave along one of
the coordinate axes of ![]() (there is a misprint in [19], where
(there is a misprint in [19], where ![]() was used, not 41.25 ). Canine ventricular conduction velocities range from
140-250 (transverse) to 500-800 (longitudinal)
was used, not 41.25 ). Canine ventricular conduction velocities range from
140-250 (transverse) to 500-800 (longitudinal) ![]() [20], so our value of D is in between longitudinal and transverse
diffusion coefficients in real anisotropic myocardium.
[20], so our value of D is in between longitudinal and transverse
diffusion coefficients in real anisotropic myocardium.
Calculations were performed using the explicit Euler method (except the `m'
gating variable which was calculated implicitly) with five-node
approximation of the Laplacian on a rectangular grid of 200 ![]() 200
to 300
200
to 300 ![]() 300 nodes with a time step of 0.01 to
300 nodes with a time step of 0.01 to ![]() and a
space step of
and a
space step of ![]() , with impermeable boundaries
, with impermeable boundaries
Spiral waves were initiated in one of three ways: by a cut wavefront,
twin pulse protocol, or a phase distribution method. A plane wave was
initiated at one edge of the medium by a ![]() duration stimulation of a
strip
duration stimulation of a
strip ![]() wide, by a current F(x,y,t) that gave a
wide, by a current F(x,y,t) that gave a ![]() of
of
![]() (which corresponds to the additional
transmembrane current of
(which corresponds to the additional
transmembrane current of ![]() ,
where
,
where ![]() is the capacitance of the membrane of one cell)
and the excitation was allowed to propagate to the
centre of the medium. The wavefront was then cut, and all the variables
on one side of the cut reset to their equilibrium values. This
numerically convenient but artificial method allows spirals to be
initiated in a
is the capacitance of the membrane of one cell)
and the excitation was allowed to propagate to the
centre of the medium. The wavefront was then cut, and all the variables
on one side of the cut reset to their equilibrium values. This
numerically convenient but artificial method allows spirals to be
initiated in a ![]() medium. The twin pulse protocol
[21], also known as the cross field technique, or
cross-stimulation [22, 23]
requires a larger (
medium. The twin pulse protocol
[21], also known as the cross field technique, or
cross-stimulation [22, 23]
requires a larger ( ![]() )
medium, in which a plane wave is initiated at the lower border by
)
medium, in which a plane wave is initiated at the lower border by
![]() stimulation of
stimulation of ![]() (
( ![]() ) of a 2 mm strip, and
) of a 2 mm strip, and
![]() later (after the wavefront has propagated through the medium,
establishing a gradient in refractoriness) the second stimulus is
applied: a
later (after the wavefront has propagated through the medium,
establishing a gradient in refractoriness) the second stimulus is
applied: a ![]() stimulation of
stimulation of ![]() (
( ![]() ) over the left
) over the left
![]() area of the medium.
area of the medium.
The phase distribution method used one-dimensional calculations to
record values of all dynamical variables in a plane periodic wave of a
high frequency, thus expressing all the 17 variables as functions of
single scalar variable, the phase. To create initial conditions, a
distribution of the phase over the plane, corresponding to an
Archimedean spiral with an appropriate wavelength, has been used to
specify the distribution of the dynamic variables via these
functions. This highly artificial method is convenient to produce
spiral wave at a prescribed location. However, it still requires large
enough medium to initiate (we used ![]() ), as the larger core
in the first few revolutions of the spiral is determined not only by the
initiation procedure, but by internal properties of the medium.
), as the larger core
in the first few revolutions of the spiral is determined not only by the
initiation procedure, but by internal properties of the medium.