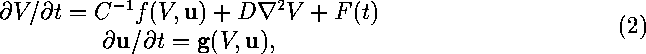The Oxsoft equations (Noble 1990) for a single isopotential cell are in the form of a system of ordinary differential equations:
where C is the capacitance of a single cell, V the membrane
potential in mV and ![]() is a vector of the activation and
inactivation gating variables and the ionic concentrations that
determine the total membrane current
is a vector of the activation and
inactivation gating variables and the ionic concentrations that
determine the total membrane current ![]() . The full
equations, together with the parameter values, are listed in the
Appendix, and can also be obtained in the form of a Pascal program in
Oxsoft HEART. This model was incorporated into a partial differential
equation model for an excitable medium in the plane (x,y),
. The full
equations, together with the parameter values, are listed in the
Appendix, and can also be obtained in the form of a Pascal program in
Oxsoft HEART. This model was incorporated into a partial differential
equation model for an excitable medium in the plane (x,y),
where D is the diffusion coefficient for V, ![]() is the
Laplacian operator
is the
Laplacian operator ![]() and F(t) is a spatially uniform, time dependent forcing that
models external electric current applied to the tissue. The diffusion
coefficient
and F(t) is a spatially uniform, time dependent forcing that
models external electric current applied to the tissue. The diffusion
coefficient ![]() was chosen to give a
conduction velocity of
was chosen to give a
conduction velocity of ![]() that corresponds to
intercellular (gap junction) conductance of
that corresponds to
intercellular (gap junction) conductance of ![]() for
for
![]() long cells (Rice et al. 1995).
long cells (Rice et al. 1995).
Calculations were performed using the explicit Euler method with
five-node approximation of the Laplacian on a rectangular grid of
200 ![]() 200 to 300
200 to 300 ![]() 300 nodes with a time and space step of
300 nodes with a time and space step of
![]() and
and ![]() (for the majority of
experiments) or 0.001 ms and 0.05 mm (i.e. less than the myocyte length) for
a few control computations. Steps used in other published ventricular
simulations were: 0.025 ms and 0.25 mm in Courtemanche & Winfree
(1991) and 0.1 ms and 0.25 mm in Efimov et al. (1995). The boundaries were modelled as impermeable,
(for the majority of
experiments) or 0.001 ms and 0.05 mm (i.e. less than the myocyte length) for
a few control computations. Steps used in other published ventricular
simulations were: 0.025 ms and 0.25 mm in Courtemanche & Winfree
(1991) and 0.1 ms and 0.25 mm in Efimov et al. (1995). The boundaries were modelled as impermeable,
with the medium large enough so that the exact form of boundary conditions does not influence the spiral wave behaviour.
At the space step of 0.1 mm, the upstroke velocity of the solitary wave
was 570 V/s, and the CV 0.38 m/s; and at space
step of 0.05 mm, 500 V/s and 0.45 m/s correspondingly.
As the space step is reduced,
the CV with ![]() converges to 0.485 m/s; the CV is
within 5% of this `true' value at
converges to 0.485 m/s; the CV is
within 5% of this `true' value at ![]() i.e. less than the
length of a single cell.
i.e. less than the
length of a single cell.
The action of external forcing as the additive term F(t) in (2) corresponds to injecting current into each cell and does not correspond to extracellular field stimulation. However, realistic models of external current require consideration of each cell as an extended object described by partial differential equations, or at least integration of transmembrane currents over each cell's surface [14] and so massively increase the computational load. This paper is mostly concerned with the spontaneous evolution of spiral waves, and the qualitative features of resonant drift and so this simplification is acceptable. We measured the external forcing F in V/s in terms of its dimensionality in (2); this can easily be rescaled to amperes per cell by dividing by cell capacitance C. The time dependent F(t) used to produce resonant drift of the spiral wave was a series of rectangular pulses of 2 ms duration, applied at a fixed time after an action potential was detected at the recording site by V increasing through -10 mV.
Spiral waves were initiated in two ways, by a cut wavefront or twin pulse
protocol. A plane wave was initiated at one edge of the medium by a 2 ms
duration stimulation of a strip 0.65 mm wide, by a current that
gave a ![]() of 50 V/s and the excitation allowed to
propagate to the centre of the medium. The wavefront was then cut, and all
the variables on one side of the cut reset to their equilibrium values. This
numerically convenient but artificial method allows spirals to be initiated
in a 2
of 50 V/s and the excitation allowed to
propagate to the centre of the medium. The wavefront was then cut, and all
the variables on one side of the cut reset to their equilibrium values. This
numerically convenient but artificial method allows spirals to be initiated
in a 2 ![]() 2 cm medium. The twin pulse protocol
requires a larger (3
2 cm medium. The twin pulse protocol
requires a larger (3 ![]() 3 cm) medium, in which a plane
wave is initiated at the lower border by 10 ms stimulation of 50 V/s
of a one mm strip, and 180 ms later (after the wavefront has
propagated through the medium, establishing a gradient in refractoriness)
the second stimulus is applied: a 4 ms stimulation of 40 V/s over
the left 3
3 cm) medium, in which a plane
wave is initiated at the lower border by 10 ms stimulation of 50 V/s
of a one mm strip, and 180 ms later (after the wavefront has
propagated through the medium, establishing a gradient in refractoriness)
the second stimulus is applied: a 4 ms stimulation of 40 V/s over
the left 3 ![]() 2.5 cm area of the medium. These large areas are
necessary for the initiation of reentry in a homogeneous tissue;
re-entry persists in smaller media, see figure 1. For anisotropic media
we took (2) with the Laplacian
replaced by
2.5 cm area of the medium. These large areas are
necessary for the initiation of reentry in a homogeneous tissue;
re-entry persists in smaller media, see figure 1. For anisotropic media
we took (2) with the Laplacian
replaced by
![]() or
or
![]() with
with ![]() ,
which is equivalent to 3-fold compression in x or y direction.
Simulating 1 s of activity in a 2
,
which is equivalent to 3-fold compression in x or y direction.
Simulating 1 s of activity in a 2 ![]() 2 cm medium with
2 cm medium with ![]() and
and ![]() took about 180 hours on a
single MIPS R8000 75 MHz central processor unit.
took about 180 hours on a
single MIPS R8000 75 MHz central processor unit.