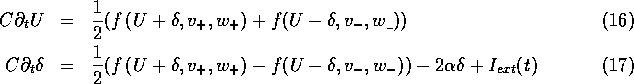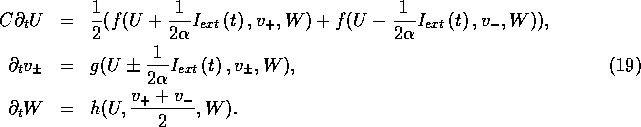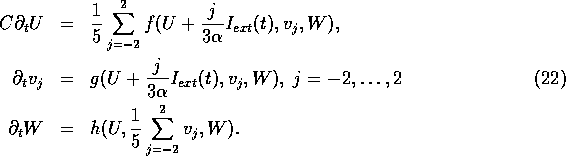The system (15) contains a singular small parameter, the
ratio of the characteristic times of the intracellular conductivity
![]() ,
, ![]() , and of the membrane excitability,
, and of the membrane excitability, ![]() .
Biophysical data [Weidmann, 1952] give the intracellular resistivity of
the order of 250 Ohm
.
Biophysical data [Weidmann, 1952] give the intracellular resistivity of
the order of 250 Ohm ![]() cm, which for the cell size of 10-80
cm, which for the cell size of 10-80 ![]() m
gives
m
gives ![]() of the order of several
of the order of several ![]() S, so
S, so ![]() , which is much less than
, which is much less than ![]() .
Numerical calculations of the simplified two-compartment model
(15) would require time steps not greater than
.
Numerical calculations of the simplified two-compartment model
(15) would require time steps not greater than ![]() , while if we can get rid of the small parameter, times steps
of around
, while if we can get rid of the small parameter, times steps
of around ![]() would be adequate.
would be adequate.
We exclude the small parameter ![]() by ``quasi-stationary'' arguments. Rewrite
the first two equations of (15) as
by ``quasi-stationary'' arguments. Rewrite
the first two equations of (15) as
where
![]()
and
![]()
Now, in (17) the term ![]() dominates over
dominates over
![]() , and omitting the nonlinear term
, and omitting the nonlinear term ![]() ,
(17) becomes a linear equation. If the characteristic time
of
,
(17) becomes a linear equation. If the characteristic time
of ![]() change is bigger than
change is bigger than ![]() , then this equation
describes the fast approach of
, then this equation
describes the fast approach of ![]() to its quasi-stationary value,
to its quasi-stationary value,
![]()
and we can substitute this value into (16), which yields
Equations (18) and the last pair of (15) form a
closed system, which depends only on the ratio ![]() . If we
assume that the duration of pulses
. If we
assume that the duration of pulses ![]() is shorter than the
characteristic time
is shorter than the
characteristic time ![]() of the slow variables then
of the slow variables then ![]() . This final simplification gives:
. This final simplification gives:
This model is almost as simple as the original (1)
ordinary differential equation (e.g., it has three equations more than
the 17 variable Noble et al. [1990] ordinary differential system we use
for ventricular excitation), but describes the effect of external
current. This has been obtained from (15) assuming the
characteristic time of the external curent pulses, ![]() , is
, is
![]()
In practice ![]() ,
, ![]() and
and ![]() are all of the order of
1 ms.
are all of the order of
1 ms.
The general model (14) can be simplified by quasi-stationary arguments to
with separate ordinary differential equations for v at each point of the
membrane. If the external field ![]() is fixed in direction and varies
only in magnitude, then the surface integral in (20)
can be reduced, in a Lebesgue style, to an ordinary integral
is fixed in direction and varies
only in magnitude, then the surface integral in (20)
can be reduced, in a Lebesgue style, to an ordinary integral
where the kernel K(s) is determined by the cell geometry, the conductivities, and the direction of the external field, and because of electroneutrality of the cell
![]()
The simple model (19) corresponds to evaluation of the integral in
(21) at two points ![]() .
.
Below we explore numerically some properties of the model (19) and its generalisation to spatially extended media. To validate the two-point approximation of the integral (21), we compare the two-point results with those of a five-point evaluation of (21), in the form